Inicio / Archivo / Año 7, No 7, septiembre 2024 - agosto 2025 / Paper 12

ACTIVIDAD ANTIFÚNGICA DE Bacillus SPP. CONTRA Fusarium
verticillioides Y Alternaria solani
Raquel Rubio-González, Israel Méndez-Meza, Lucina Hernández-Martínez, Alejandro Hernández-Morales*
Escuela de Bioquímica, Facultad de Estudios Profesionales Zona Huasteca, Universidad Autónoma de San Luis Potosí
*alejandro.hernandez@uaslp.mx
Resumen
Fusarium verticillioides y Alternaria solani son dos hongos fitopatógenos que causan enfermedades en cereales y solanáceas. Una alternativa para el control de estos fitopatógenos son las bacterias del género Bacillus que producen lipopéptidos y compuestos orgánicos volátiles que inhiben el desarrollo de hongos. El objetivo de este trabajo fue determinar la actividad inhibitoria de cinco aislados de Bacillus spp. contra F. verticillioides y A. solani. Para evaluar la inhibición, los hongos se sembraron en placas de PDA envenenadas con sobrenadante de los cultivos bacterianos; mientras que el efecto de los volátiles se determinó mediante la técnica de placa dividida. Se realizaron tres réplicas por tratamiento más un control positivo con la cepa Bacillus subtilis Q11. Además, se amplificaron los genes involucrados en la síntesis de lipopéptidos mediante la reacción de PCR. Los resultados obtenidos indican que Bacillus sp. CIS52, CIS53 y CIS54 poseen los genes involucrados en la síntesis de lipopéptidos del tipo iturinas, surfactinas y fengicinas; mientras que Bacillus sp. CIS45, iturinas y surfactinas; Bacillus sp. CIS49 fengicinas y B. subtilis Q11, iturinas; sugiriendo que pueden sintetizar al menos un tipo de lipopéptido. Aunque los aislados sintetizan lipopéptidos, solo inhibieron el desarrollo de F. verticillioides, siendo Bacillus sp. CIS49 la bacteria con mejor efecto. Respecto a los volátiles, sólo se observó inhibición de A. solani en un rango del 48.75 % al 67.73 %. Los resultados indican que F. verticillioides es inhibido por compuestos tipo lipopéptidos, mientras que A. solani es inhibido por volátiles sintetizados por Bacillus sp. En conclusión, los aislados de Bacillus spp. poseen actividad inhibitoria contra F. verticillioides y A. solani.
Palabras clave:Biocontrol, inhibición, hongos fitopatógenos.
Introducción
La actividad agrícola genera aproximadamente el 80% de los alimentos que se consumen a nivel mundial y contribuye al desarrollo económico, social y ambiental (Villarreal et al., 2018). Sin embargo, es afectada por diferentes factores ambientales entre los que se encuentran las enfermedades causadas por fitopatógenos. Las enfermedades en plantas provocan pérdidas anuales estimadas entre el 10 al 15 % en los principales cultivos del mundo, con pérdidas económicas directas de hasta cientos de miles de millones de dólares. De estas enfermedades, el 70-80% son causadas por hongos fitopatógenos.
En los últimos años, las enfermedades fúngicas de los cultivos se han vuelto cada vez más graves, ya que afectan el rendimiento y la calidad de los cultivos (Peng et al., 2021), entre estas se encuentran las patologías causadas por Fusarium verticillioides y Alternaria solani.
F. verticillioides es un patógeno necrótrofo por la capacidad que tiene de causar la muerte del tejido hospedero y sobrevive como saprófito en el rastrojo, siendo transmitido por el suelo hacia la planta hospedera, el maíz (Zea mays L.). F. verticillioides es responsable de importantes pérdidas de rendimiento al causar pudrición de raíces, tallos y mazorcas, y pérdidas en la calidad debido a la acumulación de micotoxinas en los granos (Einloft et al., 2021). Las fumonisinas son las principales toxinas que se sintetizan en el tejido del hospedero y en los granos del maíz, disminuyen la calidad e inducen toxicidad pleiotrópica en animales, incluyendo neurotoxicidad, nefrotoxicidad y hepatotoxicidad con un impacto relevante en la salud humana (Liu et al., 2019).
A. solani causa enfermedades en el follaje (tizón temprano), tallos basales de plántulas (podredumbre del cuello), tallos de plantas adultas (lesiones del tallo) y frutos (podredumbre del fruto) del tomate. El tizón temprano es la enfermedad más destructiva en especies de la familia de las solanáceas entre las cuales se encuentra el tomate, la berenjena y la papa (Chaerani y Voorrips, 2006). En las plántulas, primero se desarrollan manchas oscuras en los cotiledones, tallos y hojas verdaderas, mientras que en las hojas de la parte inferior de las plantas más viejas aparecen manchas de color marrón oscuro con anillos concéntricos oscuros. Luego se produce una defoliación severa debido a la muerte de las hojas afectadas (Muimba, 2018).
Existen diversos compuestos químicos tales como plaguicidas y fungicidas para controlar a los hongos fitopatógenos. Sin embargo, son tóxicos para el ser humano, contaminan los suelos y mantos acuíferos e incrementan el costo de la producción. Asimismo, se ha demostrado que contribuyen a la generación de resistencia en hongos fitopatógenos. Por lo que es necesario buscar alternativas sustentables para el control de los fitopatógenos fúngicos (Villarreal et al., 2018). Una alternativa sustentable es el uso de bacterias del género Bacillus, las cuales producen lipopéptidos antifúngicos como las fengicinas, surfactinas e iturinas que inhiben el desarrollo de gran variedad de hongos fitopatógenos (Harwood et al., 2018).
Adicionalmente se ha demostrado que especies pertenecientes al género Bacillus producen compuestos orgánicos volátiles entre los que se encuentran la acetoína, benzaldehído, benzotiazol y acetofenona que inhiben el desarrollo de hongos fitopatógenos (Grahovac et al., 2023). Debido a las características mencionadas, el género Bacillus spp. posee amplio potencial para el desarrollo de bioformulados con efecto antifúngico. Por lo que el propósito de esta investigación es evaluar el efecto inhibitorio de cinco aislados de Bacillus spp. contra F. verticillioides y A. solani y determinar si el efecto antifúngico es mediado por lipopéptidos o compuestos orgánicos volátiles.
Objetivo
Determinar el efecto de lipopéptidos y compuestos orgánicos volátiles de Bacillus spp. contra Fusarium verticillioides y Alternaria solani.
Materiales y métodos
Microorganismos utilizados
Los aislados de Bacillus sp. CIS45, CIS49, CIS52, CIS53 y CIS54 se recuperaron del cepario del Laboratorio de Biotecnología de la Facultad de Estudios Profesionales Zona Huasteca. Las cepas de Bacillus subtilis Q11, Fusarium verticillioides y Alternaria solani fueron donadas por la empresa SinQuimia SA de CV.
Obtención de lipopéptidos y técnica de alimento envenenado
Los aislados de Bacillus sp. CIS45, CIS49, CIS52, CIS53 y CIS54 se inocularon en 10 mL de caldo LB y se incubaron a 30 ºC con agitación a 200 rpm durante toda la noche. Una alícuota de los cultivos se transfirió a 200 mL de caldo LB y se incubó a 30 ºC con agitación a 200 rpm durante 120 h de fermentación (Cao et al., 2012). Transcurrido el tiempo de incubación, se recuperó el sobrenadante mediante centrifugación a 13000 rpm durante 30 min y se guardó en refrigeración hasta su uso. Para la técnica de alimento envenenado se utilizó la metodología propuesta por Erhonyota et al. (2023). Para ello, se pesó el agar dextrosa papa (PDA) y se disolvió en el sobrenadante recuperado de cada cultivo. El medio se esterilizó a 120 ºC, 15 lb de presión durante 15 minutos. El medio se dejó enfriar y se vertió en placas de Petri de 100 x 15 mm. Para el ensayo de inhibición, un disco de micelio de 0.5 cm de cada uno de los hongos se inoculó en el centro de las placas de PDA envenenadas. Como control positivo se utilizaron placas de PDA preparadas con el sobrenadante de B. subtilis Q11. Para el control negativo el disco de micelio de cada hongo se sembró en placas de PDA que no contenían sobrenadante. Las placas se incubaron a 30 ºC durante una semana y se monitoreó comparando el desarrollo de los tratamientos con respecto a la placa control en medio PDA. Se realizaron tres réplicas por tratamiento, incluyendo el control positivo y negativo. El análisis estadístico se realizó en el programa Graphad V10. El análisis de varianza (ANOVA) se realizó mediante la comparación de medias por el método Tukey. terminación de genes involucrados en la síntesis de lipopéptidos
Se obtuvo DNA de los cinco aislados de Bacillus sp. y B. subtilis Q11 utilizando el kit GeneJET Genomic DNA Purification K0721 (Thermo SCIENTIFIC). Se utilizaron los oligonucleótidos reportados por para amplificar los genes de las principales familias de lipopéptidos (ituD y bamC), surfactinas (srfAB) y fengicinas (fenB). Las condiciones para la reacción en cadena de la polimerasa fueron las siguientes, desnaturalización: 95 °C -10 min, 35 ciclos de 95 °C -30 segundos, 55 °C -30 segundos, 72 °C -1 min, extensión final 72 °C -5 min. Los productos obtenidos se tiñeron GelRed (Química Valaner) y se visualizaron en un transluminador Solo Elite Vision.
Evaluación del efecto de los compuestos orgánicos volátiles
La técnica de placa dividida se realizó de acuerdo con lo reportado por Kong et al. (2020). En una placa de Petri con dos divisiones se vertió agar LB y PDA de manera independiente. En el medio LB se inoculó cada una de las bacterias a evaluar, mientras que en el agar PDA se colocó un disco de micelio de F. verticillioides o A. solani. Se realizaron tres réplicas por tratamiento más el control positivo con B. subtilis Q11 y un control negativo en el cual se inoculó el disco de micelio en agar PDA, mientras que en la división con agar LB sin inoculación. El análisis estadístico se realizó en el programa Graphad V10. El análisis de varianza (ANOVA) se realizó mediante la comparación de medias por el método Tukey.
Resultados
Determinación del efecto de Bacillus spp. contra F. verticillioides
Para determinar si los aislados de Bacillus sp. CIS45, CIS49, CIS52, CIS53 y CIS54 producen lipopéptidos, se obtuvieron compuestos extracelulares sintetizados durante 120 h de fermentación en caldo LB. F. verticillioides se cultivó en las placas envenenadas y se monitoreó el desarrollo del hongo en cada uno de los tratamientos y se comparó con el crecimiento en las placas de PDA utilizadas como control negativo. Los resultados indican que Bacillus sp. CIS49 es el aislado que muestra mayor efecto inhibitorio en el desarrollo de F. verticillioides en un 34% (p < 0.05), mientras que los aislados Bacillus sp. CIS45, CIS52, CIS53, CIS54 y la cepa B. subtilis Q11 ejercen menor inhibición en un 20% aproximadamente (Figura 1).
Para determinar el efecto antagónico mediado por compuestos orgánicos volátiles los aislados Bacillus sp. CIS45, CIS49, CIS52, CIS53, CIS54 y la cepa B. subtilis Q11 se co-cultivaron con F. verticillioides, utilizando la técnica de placa dividida. Los resultados mostraron desarrollo similar de F. verticillioides en los tratamientos y en el control negativo, sugiriendo que los compuestos orgánicos volátiles producidos por los aislados de Bacillus spp. y B. subtilis SQ11 carecen de efecto inhibitorio contra el hongo (Figura 1).
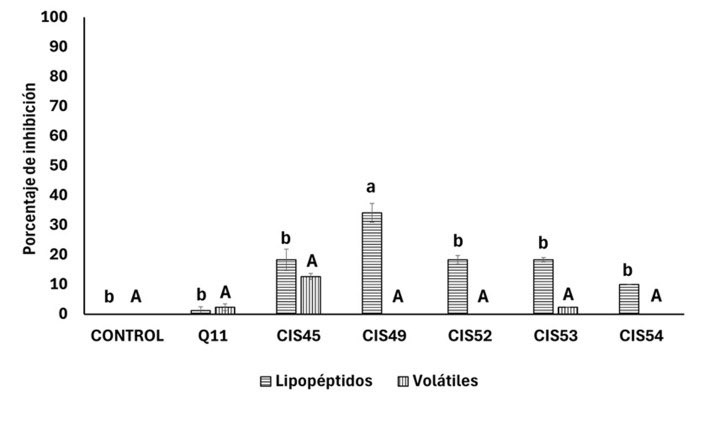
Figura 1. Efecto de lipopéptidos y compuestos orgánicos volátiles de Bacillus spp. contra Fusarium verticillioides. Nota: Letras distintas indican diferencia significativa p < 0.05.
Determinación del efecto de Bacillus spp. contra A. solani
Para determinar el efecto inhibitorio de los lipopéptidos sintetizados por Bacillus sp. CIS45, CIS49, CIS52, CIS53 y CIS54 contra A. solani, se obtuvieron compuestos extracelulares producidos durante 120 h de fermentación en caldo LB. A. solani se cultivó en las placas envenenadas y se monitoreó el desarrollo en cada uno de los tratamientos con respecto al crecimiento en las placas control. Los resultados muestran desarrollo similar de A. solani en los tratamientos y en los controles, sugiriendo que los compuestos extracelulares producidos por los cinco aislados de Bacillus spp. carecen de efecto inhibitorio contra el hongo. El análisis estadístico muestra que el desarrollo de A. solani en los tratamientos es no significativo con respecto a los controles.
En lo que respecta al efecto de los compuestos orgánicos volátiles los aislados Bacillus sp. CIS45, CIS49, CIS52, CIS53, CIS54 y la cepa B. subtilis Q11 se co-cultivaron con A. solani en placas con dos divisiones. Se observó inhibición del desarrollo de A. solani cuando se co-cultivó con los aislados y con la cepa B. subtilis Q11, con porcentajes de inhibición desde 48.75 ± 10.62% para Bacillus sp. CIS45, mientras que, para el resto de los aislados y la cepa B. subtilis Q11 se observaron porcentajes de inhibición mayores al 60%. El análisis estadístico muestra que todos los tratamientos son similares entre sí, pero diferentes al control negativo, lo cual indica que los compuestos orgánicos volátiles sintetizados por aislados Bacillus sp. CIS45, CIS49, CIS52, CIS53, CIS54 y la cepa B. subtilis Q11 inhiben el desarrollo de A. solani (Figura 2).
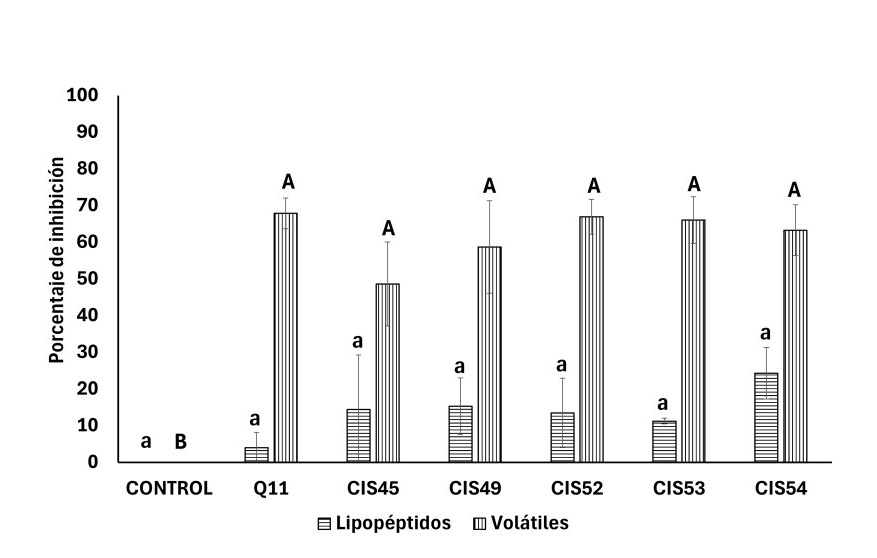
Figura 2. Efecto de lipopéptidos y compuestos orgánicos volátiles de Bacillus spp. contra Alternaria solani. Nota: Letras distintas indican diferencia significativa p < 0.05.
Identificación de los genes involucrados en la síntesis de lipopéptidos
Uno de los principales metabolitos que producen los Bacillus sp. son los lipopéptidos, los cuales han sido clasificados en tres familias iturinas, surfactinas y fengicinas. Para determinar si los Bacillus sp. CIS45, CIS49, CIS52, CIS53 y CIS54 poseen los genes para la síntesis de lipopéptidos se extrajo DNA y se amplificaron los genes involucrados en la síntesis lipopéptidos. Se amplificaron los genes bamC e ituD involucrados en la síntesis de iturinas, srfAB para la síntesis de surfactinas y fenB para las fengicinas. En la reacción de PCR se obtuvieron productos de amplificación con los tamaños esperados bamC, 850 pb; ituD, 647 pb; srfAB, 308 pb y fenB, 767 pb. En Bacillus sp. CIS45 se identificaron los genes ituD y srfAB, mientras que en Bacillus sp. CIS49 se identificó el gen fenB. Para el resto de los aislados Bacillus sp. CIS52, CIS53 y CIS54 se amplificaron los cuatro genes bamC, ituD, srfAB y fenB (Tabla 1). Los resultados sugieren que los cinco aislados podrían producir al menos un tipo de lipopéptido. En cuanto a la cepa B. subtilis Q11 solo se identificó el gen ituD.
Tabla 1. Genes identificados en Bacillus spp.
Familia |
Gen |
Q11 |
CIS45 |
CIS49 |
CIS52 |
CIS53 |
CIS54 |
|---|---|---|---|---|---|---|---|
Iturinas |
bamC |
nd |
nd |
Nd |
+ |
+ |
+ |
ituD |
+ |
+ |
Nd |
+ |
+ |
+ |
|
Surfactinas |
srfAB |
nd |
+ |
Nd |
+ |
+ |
+ |
Fengicinas |
fenB |
nd |
nd |
+ |
+ |
+ |
+ |
Discusión
El género Bacillus comprende más de 60 especies ampliamente distribuidas en la naturaleza, se encuentran en el suelo, asociados a las raíces de las plantas y se propagan hacia el agua, plantas y animales (Villarreal et al., 2018). La colonización de las raíces por bacterias del género Bacillus es benéfica para la planta huésped. Las plantas proporcionan los nutrientes, mientras que Bacillus proporciona compuestos bacterianos y actividades que estimulan el crecimiento vegetal y brindan protección contra fitopatógenos (Hashem et al., 2019). La capacidad antagónica de los Bacillus está mediada por compuestos denominados lipopéptidos y compuestos orgánicos volátiles que inhiben el desarrollo de hongos fitopatógenos (Shafi et al., 2017; Villarreal et al., 2018; Valenzuela et al., 2024). Los lipopéptidos se han clasificado en tres familias iturinas, surfactinas y fengicinas, son potentes disruptores de la integridad estructural, al digerir componentes de la envoltura celular, ya sea por lisis o inhibición de la biosíntesis de membrana. Los lipopéptidos pueden agregarse de manera selectiva, formando poros que permiten el paso descontrolado de iones y otros solutos a través de los canales, conduciendo a la muerte celular.
Para determinar si los cinco aislados de Bacillus sintetizan lipopéptidos, se amplificaron los genes involucrados en la síntesis de iturinas, surfactinas y fengicinas. Los resultados obtenidos sugieren que Bacillus sp. CIS45 sintetiza iturinas y surfactinas, Bacillus sp. CIS49 fengicinas, mientras que Bacillus sp. CIS52, CIS53 y CIS54 los tres tipos de lipopéptidos iturinas, surfactinas y fengicinas. En lo que respecta al efecto antagónico se encontró efecto moderado de Bacillus sp. CIS49 contra F. verticillioides; mientras que el resto de los aislados y la cepa B. subtilis Q11 carecen de efecto inhibitorio. Respecto a A. solani ninguno de los cinco aislados ni la cepa B. subtilis Q11 mostraron efecto inhibitorio. Los resultados difieren de los reportados previamente por otros autores quienes reportaron una fuerte inhibición de F. verticillioides por lipopéptidos de B. velezensis sintetizados durante 72 h de fermentación (Dinitz et al., 2024). Mientras que Zhang et al. (2022) reportaron la inhibición de A. solani por lipopéptidos de B. subtilis producidos durante 12 horas de fermentación. Por lo que es necesario reducir los tiempos de fermentación, analizar la expresión de los genes y comprobar la producción de los lipopéptidos por los aislados Bacillus sp. CIS45, CIS49, CIS52, CIS53 y CIS54.
En lo que respecta a la inhibición mediada por compuestos orgánicos volátiles se observó que los aislados Bacillus sp. CIS45, CIS49, CIS52, CIS53, CIS54 y la cepa B. subtilis Q11 inhiben el desarrollo de A. solani, mientras que en F. verticillioides no se observó inhibición. Se han reportado que B. subtilis, B. amyloliquefaciens y B. cereus producen compuestos orgánicos volátiles que inhiben el desarrollo de hongos fitopatógenos. En general los volátiles se clasifican en el grupo de alcoholes, cetonas y benzotiazoles (Grahovac et al., 2023). Si bien, en este estudio no se identificaron los compuestos orgánicos volátiles es probable que el efecto observado en A. solani se deba a metabolitos previamente reportados para el género Bacillus sp. Finalmente, es necesario identificar los compuestos orgánicos volátiles sintetizados por los aislados Bacillus sp. CIS45, CIS49, CIS52, CIS53 y CIS54.
Conclusión
Los bacilos estudiados inhiben el desarrollo de los hongos fitopatógenos, siendo Bacillus sp. CIS49 el aislado que muestra mayor efecto inhibitorio contra F. verticillioides, este efecto podría deberse a lipopéptidos de la familia de las fengicinas. Respecto a los volátiles, A. solani es inhibido por los compuestos orgánicos volátiles sintetizados por Bacillus sp. CIS45, CIS49, CIS52, CIS53 y CIS54. En general, los lipopéptidos inhiben el desarrollo de F. verticillioides y los compuestos orgánicos volátiles inhiben el desarrollo de A. solani.
Agradecimientos
El trabajo se realizó con el apoyo del Programa Presupuestario F003 CB2017-2018 (antes Fondo Sectorial de Investigación para la Educación CONACYT). Número de proyecto A1-S-40454. Lucina Hernández-Martínez agradece a CONAHCYT por la beca otorgada para los estudios de Maestría en Ciencias Bioquímicas con número de CVU 1230688.
Referencias
Séptimo Congreso Nacional de Tecnología 16,
17 y 18 de octubre de 2024,
celebrado en formato virtual

D. R. © UNIVERSIDAD NACIONAL AUTÓNOMA DE MÉXICO
Excepto donde se indique lo contrario esta obra está bajo una licencia Creative Commons
Atribución No comercial, No derivada, 4.0 Internacional (CC BY NC ND 4.0
INTERNACIONAL).
https://creativecommons.org/licenses/by-nc-nd/4.0/deed.es

ENTIDAD EDITORA
Facultad de Estudios Superiores Cuautitlán.
Av. Universidad 3000, Universidad Nacional Autónoma de México, C.U., Delegación Coyoacán, C.P. 04510, Ciudad de México.
FORMA SUGERIDA DE CITAR:
Rubio-González, R., Méndez-Meza, I., Hernández-Martínez, L., y Hernández-Morales, A. (2024). ACTIVIDAD ANTIFÚNGICA DE Bacillus SPP. CONTRA Fusarium verticillioides Y Alternaria solani. MEMORIAS DEL CONGRESO NACIONAL DE TECNOLOGÍA (CONATEC), Año 7, No. 7, septiembre 2024 - agosto 2025. Facultad de Estudios Superiores Cuautitlán. UNAM https://tecnicosacademicos.cuautitlan.unam.mx/CongresoTA/memorias2024/Mem2024_Paper12.html